ネオンテトラの色変化する構造色
2011年の論文
J. R. Soc. Interface 8, 56-66(2011).
に基づく解説です。
ネオンテトラの青いストライプは構造色です。この構造色には、蝶や鳥の羽根とは異なる大きな特徴があります。色が変化するのです(図1)。ストライプの青色は夜になると(周囲が暗くなると)紫色になります。明かりをつけて数分くらいすると、もとの青色戻ります。一方、餌を食べるなどして興奮すると、ストライプは黄緑色に変化することもあります。
構造色が光の干渉によって生み出されているとすれば、反射波長を決定しているのは構造の空間的な長さ(周期)です。ストライプが色変化することは、構造色を生み出している微細構造が可変であることを意味しています。
ネオンテトラの色変化はどのように実現されているのでしょうか。

ストライプの色は、虹色素胞(iridophore)と呼ばれる色素細胞から出来上がっています。皮膚表面を顕微鏡で拡大しながら観察すると、無数の青い点が集まって、ストライプ模様を形成している様子がわかります(図2)。皮膚標本を浸している生理食塩水の濃度を変えることで、構造色の色変化を誘導することができます。その様子を撮影したのが図2右上の写真です。顕微鏡下で明るく見えるこの細長い物は、虹色素胞内部にある反射小板の束であることが電子顕微鏡観察からわかっています。
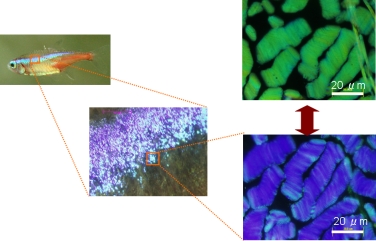
図2 ストライプを拡大していくと虹色素胞内に青い点が見える
反射小板はグアニンを主成分とする結晶でできており、板と細胞質が交互に重なった多層膜構造を形成しています。すなわち多層膜干渉による波長選択反射が青い輝点を生み出しているのです。
ネオンテトラに限らず魚一般の銀色の輝きは、グアニンの反射板によって作られていることが知られています。ネオンテトラでもストライプ以外の白い部分は、変化しない(inactive と呼ばれる)反射板が存在しています。
色が変化するには干渉を起こす多層膜構造が変わらなければなりません。干渉条件に影響するパラメータ、角度、膜厚、屈折率の中で、実際に変化できそうなのは、
角度と細胞質層の厚さです。(グアニン結晶は堅く、変化できないことが想像できます。また屈折率は材質が変わらなければ不変です。)
過去の研究から色変化の仕組みに関して、興味深いモデルが提案されていました。図3のブラインドモデルというモデルです。ちょうど窓にかかるブラインドのように、反射板の角度が変化することで細胞質の層が厚くなり、反射波長が変化するモデルです。本当にこの仕組みが正しいのか、定量的に検証するために、精密な光学測定を行いました。
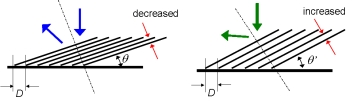

ブラインドモデルでは反射小板の角度が変化するため、光の反射方向が変わることが予想されます。この性質を実験的に確認するために、新しく顕微分光システムを構築しました(図4)。このシステムは三つの光検出器を含んでいて、虹色素胞の実像、反射スペクトル、そして反射光の方向の三つの情報を得られるようになっています。
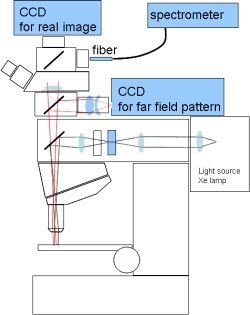
実像は顕微鏡の鏡筒上部のCCDカメラで観察します。これは通常の顕微鏡システムにあるカメラと同じで、鏡頭上部に試料の拡大像が出来ることを利用しています。二つ目の反射スペクトルは、ファイバー導入型の分光器を使って測定しています。ビームスプリッタで光を分岐し、結像面を二つ作ります。その片方に、光ファイバーの端面を配置します。最後に、光が反射される方向ですが、対物レンズの後側焦点面の中空像を観察することで、反射方向に関する情報を得ることが出来ます。この面はFar Field Patternと呼ばれ、反射光が無限遠方まで進んだときの角度依存に対応します。対物レンズに関する正弦条件により、反射方向とパターンの位置との関係が分かります。
実験では、試料を照らす落射照明が、特定の方向からだけになるように、開口絞り位置には小さいピンホールを配置する工夫を施しました。ピンホールを入れないと、対物レンズの開口全体を使った角度範囲の広い照明になってしまいます。開口絞りの像は、対物レンズの後側焦点面(光の方向を考えると前側焦点面と呼ぶべき面)にできるためレンズを通過した光はほぼ平行光線になります。このシステムには、ピンホール位置を光軸に沿って移動できる工夫が必須です。そうしないと、ピンホール像を対物レンズの後側焦点面に正しく持ってくることが出来ません。また、後側焦点面は対物レンズによって異なるので、対物レンズを交換したときには再調整が必要になります。
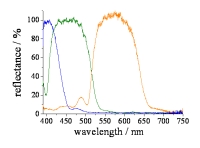
実験と解析の結果、虹色素胞の色変化(反射波長のシフト)と反射方向の変化は、ブラインドモデルでほぼ完全に説明できることが分りました。図5左は実験からえられた反射スペクトルのシフトです。生理食塩水の化学組成を変えることで、虹色素胞に色変化を誘導しました。紫外線領域の短波長から、200nm程度の長波長へのシフトが確認できます。
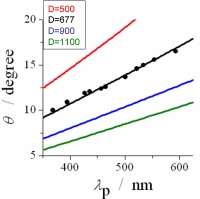
干渉条件. ブラインドモデルから予想される板の角度と反射ピーク波長の関係。
反射される光の波長と板の角度は、右の干渉条件によって関係付けられることが期待できます。この式にあらわれるパラメータを注意深く決定しました。
まず始めに、細胞質とグアニン結晶の屈折率ですが、これは既に知られているので問題ありません。次に、反射小板の厚さは原子間力顕微鏡や光分解能の表面形状を観測する顕微鏡を用いて決定しました。細胞質部分の厚さは、傾き角度に依存します。傾き角度は、光の反射方向から決定できるので、そこから逆算できます。また光の入射角度は、試料全体をゴニオメータに乗せることで制御可能にしてあります。
決定されたパラーメータから解析を行った結果が図5右です。横軸が反射光の波長、縦軸が反射板の傾き角度です。
ブラインドモデルから予想される理論曲線にぴったりのっていることが分ります。 この結果から、他の色変化に関するモデルの寄与はほとんどないことが結論できました。
それにしても、いったいどのようにして反射板の傾きを変化させているのでしょうか。虹色素胞の中に存在するモーター蛋白や反射板とつながる繊維が関係していることが分っていますが、 分子メカニズムの解明には今後も研究を続けていく必要があります。
ネオンテトラに限らず、魚やイカ・タコの仲間では、色や体の模様を変化させる性質は珍しくありません。しかも一秒以内に、赤らか青へスペクトルを変える魚の構造色が報告されています。色変化する動的な構造色に関しては、今後も研究対象に事欠くことはなさそうです。