ニシキオオツバメガの構造色:色混合効果と偏光特性
| 2007-2009年に発表した論文、
Opt. Express, 15, 2691-2701(2007). J. R. S. Interface, 5, 457-464(2008). Proc. SPIE, 7401, 740105(14pages) (2009). に基づいてニシキオオツバメガの構造色について解説します。 |
モルフォチョウを代表例として、チョウやガの仲間には驚くほど鮮やかで美しい翅を持つ種類がいます。構造色と呼ばれるこれらの色は、鱗粉の内部に存在する微細で複雑な構造が、特定の波長の光を強く反射させることで生み出されています。ちょうど、目には見えない小さな突起の配列がCDやDVD裏面の虹色を生み出したり、薄くなったシャボン玉が七色を呈するようにです。
しかし、翅の輝きの原因の全てが鱗粉の微細構造にあるわけではありません。チョウの工夫はもっと複雑で、微細な構造体だけでなく、色素の併用、鱗粉の重複など、
軽くて薄い翅の上には数多くの工夫が集積され、総合的な方法で美しい色が実現されているのです。強い湾曲を利用したニシキオオツバメガの構造色はその代表例といえるでしょう。ここではそのことを明らかにした論文に基づいて、その仕組みを紹介します。


オオツバメガ亜科(Uraniinae)に属する昼行性のガには、黒地に鮮やかなストライプ模様を持つ種類がいます。例えば、マダガスカルに生息するニシキオオツバメガや、長距離の渡りで有名なナンベイツバメガ(Urania fulgens)です。これらのガは、モルフォチョウと並んで構造色の代表例とされ、二十世紀中ごろの研究論文には、チョウの構造色をMorpho typeとUrania typeの二つに分類してあるものも見つけることができます。
顕微鏡で翅をのぞくと、ニシキオオツバメガの構造色の奇妙さがすぐにわかります。近縁種であるナンベイツバメガとの比較をしながら話を進めましょう。図2の上に示すように、ナンベイツバメガの翅には、金属のように輝く鱗粉が並んでいます。この鱗粉は上層鱗と呼ばれ、上下に二層ある鱗粉配列のうち、上側に露出した鱗粉です。下層鱗は真っ黒な色をしていて、上層鱗の下を裏打ちして色を鮮やかにしています。
下層鱗一枚の形は写真では黒いためにわかりませんが、
金属光沢のある上層鱗は、少し間隔をあけながら配列しているので、一枚の形状がはっきりと分かります。
一方、図2下に示すニシキオオツバメガの鱗粉配列は奇妙です。まず、鱗粉がほぼ隙間なく配列しているために、鱗粉の輪郭がはっきりとわかりません。また、配列した鱗粉の複数に広がって、明るい横帯の縞模様が見えます。しかもその横帯には、細い帯と太い帯の二本があります。この太さの異なる二本の帯は、色もはっきりと違って見えます。


の鱗粉配列写真(光学顕微鏡写真)。
二色の横帯が奇妙である。


構造をより詳しく比較するために、走査型電子顕微鏡を用いて観察すると、鱗粉の湾曲形状に大きな違いがあることが分かりました。ナンベイツバメガの鱗粉は、ほぼ平らに写っていますが、 ニシキオオツバメガの鱗粉はに示すように、縦方向にくるりと湾曲をした形状を持っているのです。鱗粉は先端方向に行くにつれて下側にカーブし、隣の鱗粉の上に重なっています。 このため、縦方向に隣接する鱗粉の間は、谷間のように凹んだ形状を持っています。
発色の直接的な原因になるのは、光の波長よりもさらに小さな構造なので、
さらに倍率を上げて透過型電子顕微鏡で観察を行いました。図4はニシキオオツバメガの鱗粉の断面の極めて小さな領域を観察したものです。湾曲した鱗粉底部の板は、実は複数のクチクラの膜で出来上がっています。膜と膜の間は大部分が空気ですが、クチクラの小さなブロックによって間隔が保持されています。
このように、異なる材質の膜が複数積み重なった構造は多層膜構造と呼ばれ、構造色を生み出す典型的な構造です。ナンベイツバメガにも同様な多層膜構造が報告されており、鱗粉の輝きを生み出す直接的な構造は、両者で共通であるといえます。多層膜構造は、ほとんど100%の割合で光を反射することができるため、レーザーを反射させる誘電体多層膜ミラーにも応用されています。

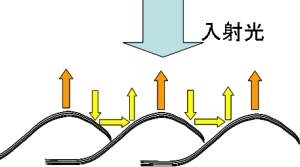
二種類のツバメガを比較してきましたが、どちらの鱗粉も空気とクチクラの多層膜構造をもち、それよって構造色を生み出していることには変わりありません。しかし、鱗粉の配列を観察すると図2のように大きく異なっているのです。それには図3で見られた鱗粉全体の大きな構造(湾曲)が深く関係しています。
図5に示す反射モデルを考えるとニシキオオツバメガの奇妙な帯の模様をうまく説明することがでます。モデルの特徴は、二つの光反射経路があることです。一つは湾曲の頂上付近から直接反射される経路、もう一つは横方向に反射された光が、隣の鱗粉でもう一度反射されて上方向に向かう二重反射経路です。実際、モデルに対応するように、二本の明るい帯は鱗粉の頂上付近と谷間の部分に観察されています。また、二重反射経路では、多層膜構造に対して斜めに(およそ45度で)光が入射するので、角度による干渉条件の違いが細い帯と太い帯の色の差を説明することができます。
図6に示すように、二色の帯模様は翅の広い範囲で見ることができます。
しかし、二本の帯は、鱗粉一枚くらいの長さ、およそ0.2mm程度しか離れていません。顕微鏡では、はっきりと分離できますが、裸眼で観察すると二色が混合された色として翅の色が見えてしまいます。
このような効果は色混合効果とよばれ、
赤緑青の三原色の輝点の強度比で色を表現するカラーテレビの原理そのものです。
点描で中間色を構成する印象派画家の作風にも似ているかもしれません。
このような色混合を用いた発色の仕組みは、自然界では特殊例のように思われるかもしれません。しかし、ニシキオオツバメガのほかにも、アゲハチョウやシジミチョウの仲間、さらにはハンミョウなどの甲虫、鳥の羽根でも発見されています。
自然界の生物はなんと複雑なことをやってのけるのでしょう。


湾曲した鱗粉の形状が生み出す色混合効果を持つ動物の例は複数ありますが、
このガの色混合は強い偏光依存性を示すのが他には見られない特色です(図7)。
詳しい理屈はここでは省略しますが、一般に多層膜構造に斜めに光が入射する場合、その反射率は偏光方向に大きく依存しています。(フレネルの公式で分かるように、S波に比べてP波の反射率が弱い)そのために、斜め入射を二度経る、二重反射経路の光は偏光に依存して大きく強度を変えるのです。
混合されるべき二本の帯のうち、片方の強度が偏光に依存するため、結果として翅全体の色も偏光によって変化することになります。無偏光の光で照明していても、見る側の方に偏光選択性があるだけで、色が変わって見えるのです。
人間にはほとんど区別のつかない光の偏光ですが、昆虫一般の眼は偏光を区別することが出来るといわれています。偏光によって変化する色には、なんらかのメッセージがあるのかもしれません。(わずかに存在する人間の眼の偏光特性には、”ハイディンガーのブラシ”が良く知られています。)
このガの構造色に関連しては、他には
・色混合効果を利用した偏光依存する三原色基板のデザイン
・高次の干渉が関与した発色の原理と色模様
・二重反射経路が示すレトロリフレクション
・顕微分光を利用した色混合の定量的評価
などについての研究も行いました。