青い翅の真白なストライプ:モルフォキプリスの構造色
|
2006年に発表した論文
Proc. R. Soc. Lond. B, 273, 129-134(2006). J. Opt. Soc. Am. A, 23, 134-141 (2006). に基づく解説です。 |
チョウの翅の色模様は、異なる色の鱗粉が並ぶことでモザイク模様のようにできあがっています。例えば、図1のミヤマカラスアゲハでは、赤い鱗粉が並んだ回りに青い鱗粉が配列することで円弧状の模様になっていますし、黒い部分にキラキラと写る輝点は、構造色の明るい鱗粉が散在してできています。ジャノメチョウの目玉模様は、同心円状に白、黒、黄色の鱗粉が並ぶことで作られています。


青さが特徴的なモルフォチョウは、モルフォ属に分類されるチョウの仲間です。 種類の数は少なくとも数十はあるようですが、その中にはストライプ模様の翅を持つ種類がいます。キプリスモルフォ(Morpho cypris)はその一種で、真っ白なストライプと青い翅が、美しいコントラストを生み出しています。そして、その模様を形成しているのは、青い鱗粉と白い鱗粉の配列です。鱗粉の青さは内部の微細構造に原因があることが分かっています。一方の白い鱗粉はどのような構造を持っているのでしょう。青の鱗粉と白の鱗粉では何が違うのでしょうか。

翅の上には青い鱗粉と白い鱗粉が並び、ストライプ模様を作る。
翅の”白さ”にも疑問があります。一般に、白い色は、光が波長依存性なく多重に散乱されることによって生み出されます。雲や雪が明るく白いのは、水滴や水の結晶が厚く集まっているために、内部で光を何度も散乱・反射し、その結果波長依存性なく反射率が上昇するからです。一方、蝶の翅は非常に薄く、鱗粉一枚の厚さは数ミクロン程度です。翅の両面にある鱗粉と翅の膜をあわせても10ミクロンには達しません。そのような薄さで、どのようにして、真白なストライプを生み出すのでしょうか。反射率が低い灰色になるべきではないでしょうか。
これら疑問に答えるために、真っ白なストライプを持つモルフォキプリスを題材に研究を行いました。その内容を以下で紹介します。


下は走査電顕写真。左は青、右は白の鱗粉断面。左上:青色が強く見えない照明配置、右上:白い鱗粉も照明の方向を変えると青い輝きをみせる角度がある。
光学顕微鏡で翅を観察すると、ストライプ模様を形成する青い鱗粉と白い鱗粉がわかります。さらに、翅を照明する光の方向を変えながら詳しく観察すると、
白い鱗粉も、青色に輝くような角度があることに気がつきました(図2右上)。
青い鱗粉の断面を走査型電子顕微鏡で観察すると、モルフォチョウに特有の棚型の構造がびっしりと林立した鱗粉構造が見られます(図2下)。意外なことに、白い鱗粉にも、同じような微細構造が観察されました。構造のスケールや形を詳しく調べましたが、大きな違いを二種類の鱗粉に見つけることは出来ませんでした。
それでは、何が青と白の色の差を生み出しているのでしょうか。鱗粉一枚で徹底的に光学測定を行うことにしました。実は、鱗粉一枚で定量的に反射率・透過率を測定するのは簡単なことではありません。鱗粉が小さく(100ミクロン程度)、しかも反射光・透過光が拡散的にひろがってしまうので、全量を測定することが難しいのです。そこで、図3のような積分球を用いた光学系を構築し、絶対反射率と透過率を測定しました。簡単に説明すると、キセノンランプからの白色光をモノクロメータを用いて単色化し、一度ピンホールに集光します。キセノンランプを用いるのは、スペクトル帯域が可視光全域に広がっていること、輝度が高く一点に集光しやすいことからです。ピンホールの大きさは十分小さく、その像をカメラ(現像)レンズで鱗粉の上に作り出します。光学系に積分球を含めることで、拡散的な光の全量(に比例する量)を検出することができます。光は光電子増倍管で検出し、ロックインアンプで増幅するシステムになっています。
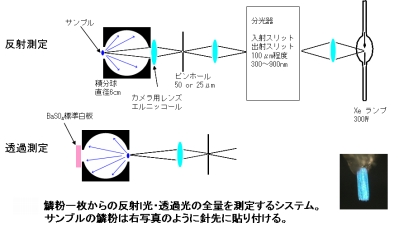
鱗粉に入射した光のエネルギーは、反射される部分、透過する部分、そして吸収されてしまう部分、の三つに分かれていきます(蛍光が無視できることは確認しました)。積分球を用いた測定から、その三つの割合をグラフ化したものが図4です。透過と反射が実験で測定できるので、残りを吸収に割り当てています。
青と白の鱗粉の差は、光の吸収量にはっきりと表れました。青い鱗粉には、吸収がかなりの割合であるのですが、白い鱗粉にはほとんどそれがありません。吸収は鱗粉が持つメラニン色素によるものと考えられますので、その含有量が二つの鱗粉ではでは大きく異なっていることを意味します。
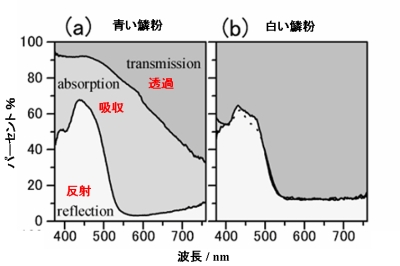
一方、短波長側(500nm以下)での強い反射は共通です。この反射は鱗粉内部の棚型の微細構造によるものと考えられますので、白い鱗粉にもその構造が観察されたことと合致しています。棚型の構造は、異方的に光を反射するので、照明光と観察角度が
ちょうど良い条件になると、白い鱗粉でも青い輝きをみることができるのです(図2右上)。
光の吸収量が大きく異なることがわかりましたが、このことがストライプの白さを説明しているわけではありません。観察者は、光の吸収量を見ているわけではなく、反射した量を見ているからです。その反射率に注目すると、白い鱗粉で、緑や赤に対応する530nm以上では、たったの13%程度しかありません。このくらいの反射率でストライプが真白に見えるのでしょうか?
実は、鱗粉だけを見ていては、翅の白さを説明することはできません。
さらに研究を進めていくと、ストライプの白さには翅全体での反射機構が関係していることが分かりました。
キプリスモルフォの翅をあらためて観察したのが図5です。これまでは背側の青地に白いストライプ模様に注目してきましたが、じつは腹側にも茶色と白の鱗粉でできたストライプがあります。さらに、鱗粉を鱗粉転写法を用いてはがして観察すると、翅の膜にも茶色と透明な部分でストライプが形成されていることが分かりました。ストライプ模様は、表面の鱗粉パターンだけではなく、裏面や膜のパターンにまで対応しているのです。

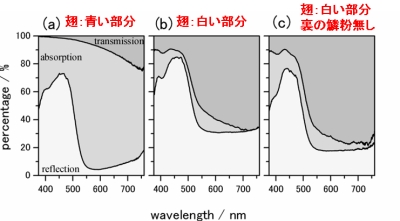
鱗粉一枚ではなく、翅そのままで積分球を用いた測定を行い、光学特性を評価すると、図6のようになりました。翅全体で測定したストライプ部分(図6中央)は、長波長側でも反射率が上昇し30%以上になっています。鱗粉一枚と比べると二倍以上の反射率です。また、裏面の鱗粉を取り除いて実験すると、反射率が低下し、翅全体の反射率には、直接光があたらない裏側の鱗粉も、確かに寄与していることが分かりました。
反射率の上昇を定量的に解析するため、翅の構造を三層モデルとしたのが図7です。モデルの詳しい説明は省略しますが、三層の間での多重反射を考慮しています。 実験により、三つ層のそれぞれがもつ反射率を注意深く決定しました。 その値を用いてモデル計算を行ったのが図8です。三つの層が関与して反射率が 高くなっていることが確かめられました。
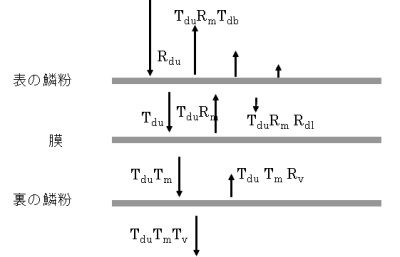
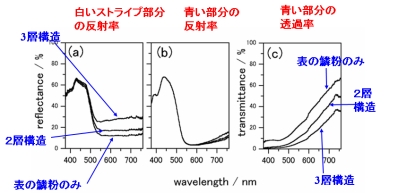
図7 翅の三層モデル(上)とモデルから
予想される反射率と透過率(下)
白さの比較のために、手元にあった普通の白い紙(コピーペーパー)で反射率を測定しました。その結果およそ70%の反射率が得られました。それに比べると、チョウの翅の30%という反射率は必ずしも高い数字には思えないかもしれません。しかし、翅はせいぜい10ミクロンの厚さなのに対して、コピー用紙の厚さは100ミクロンくらいです。仮に、厚さで規格化した反射率を考えるなら、チョウの翅の方がずっと効率が良いことになります。
チョウの翅は飛ぶための大切な道具ですから、重くすることは出来ません。
一方、白さを出すためには、材質は厚くする方が有利です。この相反する要求を満たすために、モルフォキプリスは翅の構造の最大限利用しているのです。
翅が鱗粉のなかで形成されるとき、袋状の構造として最初は作られます。そのため表面、裏面の鱗粉模様は基本的には空間的に離れたプロセスで発生します。その模様がきっちり三つの層で対応しているあたりに、生物のもつ奥深さ・不思議さを感じてしまいます。